
研究
痛み・心理社会的要因
2026年5月29日
痛みをコントロールする免疫機序:基礎となる生理学的メカニズムを探り、理論的な考察から理学療法アプローチを導くために
はじめに
理学療法士は、未解決の痛みを抱える患者さんに対応する中で、気持ちが圧倒されてしまうことがあるかもしれません。さらに、保存的治療が患者さんの訴えに与える効果は、やや限定的なことも多いです。 痛みと機能障害の理解は、主に pain drivers and disability model に基づいています。これは、さまざまな関連要因をより正確に捉えることを可能にしました。 認知的・情動的なドライバーが痛みの治療アプローチとして注目されるようになってきたことで、特に慢性疼痛の文脈では、臨床家が侵害受容(nociceptive)の領域を見過ごしてしまうことにつながった可能性があります。 このレビューでは、理学療法士が免疫による痛みの制御(immune control of pain)について、アクセスしやすい基礎知識を得られるようにすることを目的としています。臨床理解と視点を広げるためです。
方法
このナラティブ・レビューは、国際的な査読付きジャーナル『Joint Bone Spine』に掲載されました。
結果
痛みの分類
侵害受容性疼痛は、局所の組織環境の変化に反応して、末梢の侵害受容器が活性化されることで生じます。 侵害受容器は、トランジエント受容体電位(TRP)チャネル、Gタンパク質共役受容体(GPCR)、電位依存性ナトリウムチャネルなど、さまざまな分子センサーを発現しており、機械的刺激・化学的刺激・熱刺激を検知します。 一度活性化されると、電気的信号を中枢神経系へ伝達します。
免疫細胞は、特定の慢性疾患において炎症性シグナルの持続に関与し、侵害受容器(ノシセプター)の感作を長引かせることがあります。 関節リウマチのような疾患では、明らかな炎症活動が低下しても痛みが持続することがあり、痛みの免疫による制御と侵害受容のシグナル伝達の複雑な相互作用が示唆されます。
神経障害性疼痛は、体性感覚の神経系における病変または疾患によって、その機能が変化することで生じます。 よく見られる疾患には、多発性硬化症や糖尿病性神経障害などがあります。 アロディニア(通常は痛みを起こさない刺激によって誘発される痛み)や、痛覚過敏(有害な刺激に対する痛みの反応が過度に大きくなる状態)が特徴として挙げられます。 神経障害性疼痛は、影響を受けた神経に持続的な構造変化と分子レベルの変化が起こることで、慢性化することがあります。
ノシプラスティックペインは、組織損傷の明確な根拠や体性感覚系の病変がないにもかかわらず、侵害受容(ノシセプション)の処理が変化して生じる痛みとして定義されています。 この論文では、この痛みの表現型についてさらに詳しくは述べていません。 線維筋痛症や、慢性腰痛の一部の病態などは、ノシプラスティックな痛みのメカニズムと関連していることがよくあります。
この論文は、痛みの免疫による制御への貢献 痛みは長い間、主に神経のプロセスだと考えられてきました。しかし、免疫メカニズムがその調節において重要な役割を果たすことを示すエビデンスが増えています。
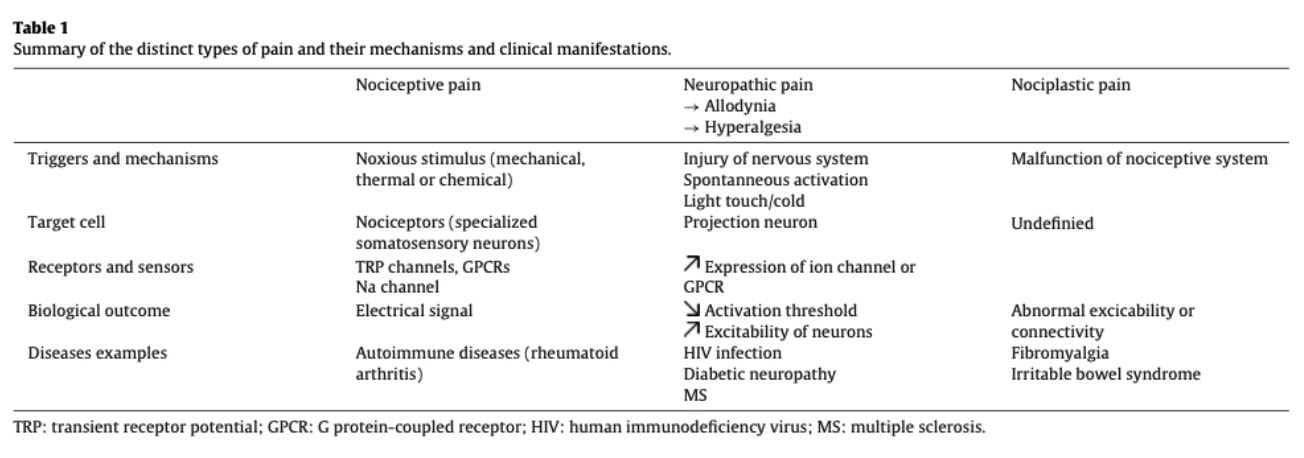
痛みを調整する免疫細胞
さまざまな免疫細胞のタイプが、痛みのメカニズムに関与していることが示されています。
マクロファージは、活性化状態のスペクトラムを取り得る非常に可塑性の高い免疫細胞で、しばしば「炎症性」と「修復(促進)性」のフェノタイプ(M1-様、M2-様)に大まかに整理されます。 組織損傷の後、マクロファージはIL-1β、TNF-α、IL-6、そしてケモカインなどのメディエーターを放出し、侵害受容器(ノシセプター)の感作や炎症性疼痛に関与します。
解決(リゾリューション)期には、マクロファージがプロ解決型の表現型へとシフトし、炎症シグナルを抑えて組織の修復を促します。 さらに、内因性オピオイドペプチドを放出し、侵害受容器にあるオピオイド受容体を活性化することで、末梢性鎮痛に寄与します。 加えて、マクロファージ由来のIL-10が炎症を制限し、痛みの解決(鎮静)をサポートします。
マクロファージと侵害受容器は、双方向のコミュニケーションを行います。 CGRP、サブスタンスP、CCL2を含むケモカインなど、侵害受容器由来のメディエーターは、マクロファージの動員や活性化状態を調節できます。 この神経免疫クロストークは、炎症の状況によって痛みを増幅もしくは鎮静化させ得ます。そして、その調節不全は慢性痛が持続することに関与していると考えられています。
ミクログリア は、中枢神経系の常在免疫細胞であり、しばしば「CNS常在のマクロファージ様細胞」として説明されます。 神経系の損傷やストレスが起きると、細胞外ATPが放出され、ミクログリア上の P2X4 や P2X7 のようなプリン作動性受容体を活性化します。
この活性化は、脳由来神経栄養因子(BDNF)を含むメディエーターのミクログリアからの放出を促し、脊髄の投射ニューロンに作用して興奮性を調節します。 これは、痛みの経路におけるニューロンの感作に寄与し、神経障害性疼痛の発症と維持において重要な役割を果たします。
T細胞は、痛みの重要な調節因子として注目が高まっている適応免疫細胞です。 異なるサブセットがそれぞれ異なる役割を担い、特にCD4+ Tヘルパー細胞は、IFN-γやIL-17などのサイトカインを産生することで、また顆粒球系のメディエーターとしてはグランザイムやパーフォリンなどを介して痛みに関与し得ます。 IL-17は機械的痛覚過敏を促進することが示されており、動物モデルでは中和によって痛みの感受性が低下します。これは、関節炎のような炎症性疾患におけるIL-17の関連性を裏付けています。
CD4+ T細胞は、プロノシセプティブな作用だけでなく、痛みの調整にも関与しています。T細胞の欠乏によって、オピオイドを介した痛みのコントロールを含む内因性の鎮痛メカニズムが損なわれるためです。
T細胞は、神経ペプチドに対する受容体を発現することで、双方向の神経免疫コミュニケーションにも関与します。 サブスタンスPやCGRPのような神経ペプチドは、T細胞の分化に影響を与え、Th17のような親炎症性の表現型を促進します。これにより、痛みの感作に寄与します。
B細胞は抗体産生細胞(免疫グロブリン)です。 免疫グロブリンG(IgG)と呼ばれる特定のB細胞が、神経障害性疼痛に関与している可能性があります。 免疫グロブリンGのレベルが上昇していることが、マウスの後根および慢性疼痛患者で確認されました。 一方で、免疫グロブリンGを枯渇させると、マウスでアロディニアはすべて防げました。 免疫グロブリンG(IgG)は、侵害受容性求心線維に発現しているFcγ受容体シグナル伝達を介して、侵害受容の感作に寄与し得ます。 この機序は、炎症性・自己免疫性の状態で痛覚過敏を高める可能性があり、さらに一部の実験モデルでは、明らかな組織損傷とは無関係に痛みに関与し得ます。 そのため、抗体を介した免疫メカニズムを標的にすることは、関節リウマチのような疾患で有望な戦略になり得ます。
好中球は自然免疫系の早期対応者で、炎症性が非常に高いです。 痛みにおける役割は状況(文脈)次第です。 急性期の痛みでは、好中球の枯渇が痛覚過敏に影響しないことが多く、急性の痛みは主に、直接的な侵害受容(ノシセプター)の活性化と、初期の炎症メディエーターによって引き起こされていることを示唆します。 慢性疼痛モデルでは、好中球が感覚神経節(例えば、脊髄後根神経節)へ浸潤し、痛覚過敏の維持に関与している可能性があります。 異なる好中球の表現型は、それぞれ異なる機能的な影響を持つと考えられます。 臨床的には、急性の腰痛から回復した患者では、慢性疼痛を発症する患者と比べて、循環中の好中球が早期に増加することが報告されています。これは、痛みの解消における初期の免疫反応の役割があり得ることを示しています。
ナチュラルキラー(NK)細胞は、自然免疫系の一部です。 その役割は、異常または損傷した細胞を検出し、排除へと導くことです。 坐骨神経痛への関与が示されています。軸索の損傷後に、神経環境内にデブリ(残骸)が蓄積し、NK細胞がそれをクリアすることで、健康的な再生環境の形成を促進するのです。 また、NK細胞は、過興奮の状態にあるニューロン、機能不全の支持細胞、持続する炎症ドライバーを標的とすることで、調節的な役割を担う可能性もあります。 これらの知見は、NK細胞が持続性疼痛の状態を理解するうえで、さらにはその調整(モジュレーション)においても関心の対象になり得ることを示唆しています。
肥満細胞は末梢神経の終末の近くに存在し、ヒスタミン、サイトカイン、プロテアーゼなどの炎症促進性メディエーターを放出することで、痛みの調節に関与します。 これらの物質は侵害受容器を活性化し、痛みの経路の感作を促進します。 また、肥満細胞は自然免疫系の細胞を呼び寄せることで、炎症カスケードを増幅し、さらに末梢の感作を強めます。
末梢性の感作
末梢性感作とは、組織の損傷や炎症の後に、侵害受容器(ノシセプター)が反応しやすくなり、作動閾値が低下することを指します。 傷ついた組織の細胞、常在の免疫細胞、さらに動員された免疫細胞は、TNF-α、IL-1β、IL-6、プロスタグランジン、ケモカインなどの炎症性メディエーターを放出します。これらは、侵害受容器に直接または間接的に作用して興奮性を高め、痛みのシグナルを増幅します。
神経免疫経路
免疫細胞は、IL-1β、IL-6、IL-17、IFN-γ、プロスタグランジンE2(PGE2)、ヒスタミン、ケモカインなどのサイトカインや炎症性メディエーターを分泌することで、侵害受容器(ノシセプター)の活動を調節します。 一方で、神経系は、CGRPやサブスタンスPといった神経ペプチドの放出によって免疫反応を調節でき、これらは免疫細胞の動員や活性化に影響を与えます。 この双方向のコミュニケーションは、痛みの増幅または解消に関わります。
中枢性感作(セントラル・センシティジーション
末梢からの侵害受容性の刺激が持続すると、中枢神経系で過剰な興奮状態(感受性亢進)が生じることがあります。特に、脊髄の後角ニューロンや、痛みを処理する上位中枢(supraspinal)の領域で起こりやすいです。 この状態は「中枢性感作(central sensitization)」と呼ばれ、痛みの感受性の増幅、アロディニア(痛みではない刺激が痛みとして感じられる状態)、広範囲に及ぶ痛み、そして組織が治癒した後でも続く持続痛と関連しています。
中枢性感作への免疫の関与
免疫細胞は、ミクログリアの活性化、サイトカインの放出、さらには神経のイオンチャネルや受容体への調節といったメカニズムを介して、中枢性感作に関与します。 例えば、ミクログリアはP2X4やP2X7のようなプリン作動性受容体を通じて細胞外ATPを感知し、その結果、脳由来神経栄養因子(BDNF)を含むメディエーターが放出されます。これにより、ニューロンの興奮性が高まります。 慢性痛は、視床下部―下垂体―副腎(HPA)軸や交感神経系の活性化を通じて免疫機能を変化させることもあり、その結果、免疫応答の調節不全や、内因性の痛みの調節能の低下につながる可能性があります。
免疫応答
免疫感受(immunoception)とは、中枢神経系が免疫の働きをモニタリングし、調整できる能力を指します。 ニューロンは、TNFRのようなサイトカイン受容体や、TLR4のようなパターン認識受容体など、炎症性シグナルを検知できる受容体を発現しています。 これらの仕組みにより、中枢神経系は免疫の活性化を検知し、それに応じて行動、代謝、そして生理学的な反応を適応させることができます。
免疫グラム
免疫グラム(immunengram)とは、過去の免疫状態の神経学的表象、または記憶痕跡として提案されているものを指し、中枢神経系と末梢の免疫組織の間で分散して存在すると考えられます。 この考え方では、免疫の経験は免疫細胞の中だけでなく、神経—免疫コミュニケーションに関与する神経ネットワークの中にも符号化されうることが示唆されています。 こうした新たなパラダイムは、神経活動が免疫の調節にどのように影響しているのか、そして自己免疫疾患のプロセスにどう関わり得るのかを説明する手がかりになる可能性があります。
免疫の自己調整と痛みの解消
多くの免疫細胞は痛みの感作や侵害受容の調節に関与しますが、免疫システムには炎症と痛みを収束させるための内因性の調節メカニズムも備わっています。 そのため、免疫細胞は生物学的な状況、タイミング、そして微小環境に応じて、侵害受容を促進する作用にも、抑制する作用にも働き得ます。
急性痛覚(ノシセプション)における免疫の関与
侵害受容性疼痛は速やかに起こり、主に神経の活性化によって媒介される一方で、免疫由来のシグナルも侵害受容器(ノシセプター)の調節に関与しています。 神経成長因子(NGF)のような分子は、免疫細胞や組織細胞によって産生され、活性化のしきい値や興奮性を調節することで、侵害受容器に対して持続的な感作(トニックなセンシタイジング)作用を及ぼします。 NGFが欠如すると、痛みを伴う刺激への感受性が低下します。これにより、急性の侵害受容においても免疫シグナルが寄与していることが強調されます。 腫瘍壊死因子α(TNF-α)のシグナル伝達も、侵害受容性の調節に関与しています。 TNF-αのシグナル伝達がない場合、侵害受容器はNGFに対する感受性が高まり、さらに異常な軸索の伸長が起こります。その結果、痛みの応答が増強されます。
侵害受容性(痛みの感知)に関わる免疫メディエーター(賛成/反対の両方
免疫細胞は多数のサイトカインやメディエーターを放出し、それが侵害受容器の興奮性や痛みの調節に直接影響します。 IL-1β、IL-6、TNF-α、IL-17のような炎症促進性サイトカインは、侵害受容器の感作や炎症性疼痛を促します。一方で、IL-10、IL-4、TGF-βを含む抗炎症性サイトカインは鎮痛作用を発揮します。 IL-10は侵害受容器に直接作用して痛みのシグナル伝達を抑えますが、IL-4は侵害受容器の感作を抑制し、マクロファージによる内因性オピオイド産生を促進します
制御性T細胞と慢性疼痛の移行
急性期から慢性期への疼痛の移行は、免疫の調節メカニズム—特に制御性T細胞(Treg)—が一部関与している可能性があります。 Tregは、エフェクター免疫細胞の働きを抑え、抗炎症性サイトカインを分泌することで、過剰な炎症反応を抑制します。 坐骨神経損傷の実験モデルでは、低用量IL-2の投与によりアロディニアが軽減し、Tregを介した鎮痛作用が示唆されました。 同様に、TNFR2アゴニストは、神経の損傷を抑え、末梢および中枢の炎症を弱めるとともに、中枢神経系内で修復に向けた免疫表現型を促進することが示されています。 これらの結果は、免疫調節が疼痛の慢性化を抑える上で重要な役割を担っていることを支持しています。
P2X7受容体と神経障害性疼痛
P2X7受容体(P2X7R)は、ATPにより開くイオンチャネルで、免疫細胞に発現しています。炎症性疼痛と神経障害性疼痛のメカニズムに重要な役割を果たします。 P2X7Rの活性化は、炎症性シグナル伝達を促進し、特にIL-1βの放出を増やします。 P2X7Rの発現増加とIL-1βレベルの上昇は、神経障害性疼痛患者で観察されており、この経路が慢性疼痛の病態生理に関与している可能性が示唆されます。 これらの結果は、痛みの感作と持続における神経免疫シグナル伝達の重要性を、さらに裏付けるものです。
鎮痛許容型の微小環境
直接の侵害受容器の調整に加えて、免疫細胞は他の免疫細胞との相互作用や、組織の微小環境を形作ることで、痛みの寛解に関与する可能性があります。 特定の免疫フェノタイプは、抗炎症・修復(reparative)のプロセスを促進し、鎮痛が起こりやすい環境(analgesia-permissive)を作り出すことで、回復を助け、慢性的な感作を抑えます。 そのため、免疫細胞と感覚ニューロンの動的な相互作用は、痛みの増幅と寛解の両方において基本的な要素であるように思われます。
質問と感想
痛みの根本的な病態生理メカニズムを理解することは、理学療法士が痛みのマネジメントに対する視点を広げるために欠かせません。 痛みのディスアビリティ&ドライバーモデルによれば、侵害受容性および神経障害性のメカニズムに関する臨床家の理解は、予後と治療の方向性の両方を見通す上で重要です。 侵害受容性および神経障害性の痛みのドライバーを特定するのに役立つ、さらなる臨床ツールについては「Talk nerdy to me」のセクションで扱い、これまでのレビューで提案されている 痛みの免疫制御とどう関連し得るのかを探ります。
生物学的に根拠づけられたモデルは、人の身体を仕組み(メカニズム)的に捉える考え方に基づいています。つまり、生理学的な経路を変えることで痛みが軽減し、機能が改善すると想定するわけです。 この視点は痛みの科学に大きく貢献してきましたが、それでも慢性痛の複雑さを十分に説明し切るには、なお不十分な面があるかもしれません。 痛みは、身体に宿る(身体的な)かつ主観的な体験であり、複雑で動的なシステムから生じます。このシステムでは、生物学的・心理的・状況(文脈)的要因が相互に作用しており、意味のある形で「孤立したサブシステム」に切り分けて説明することはできません。
ヘルスサイエンスにおける現象学は、個人が体験している「生の経験」に焦点を当てることで、このより広い理解に貢献します。 著者が定義しているとおり: 「現象学とは、体験している当事者の意識に基づいて、その体験に付与されている意味を観察し、記述しようとする哲学的潮流である」(https://doi.org/10.3917/rsi.081.0021)。 この枠組みの中では、理学療法の実践は、とりわけ統合的で患者中心のアプローチを取り入れるのに適しています。 臨床場面における反復的なやり取りによって、患者の経験を段階的に掘り下げていくことができ、意味のある治療方針を一緒に作り上げていく(共同構築する)ことが可能になります。
この文脈において、理学療法士は単に不調を「治す」ことを目的とした技術者にとどまるのではなく、患者と協働していく臨床パートナーです。ケアを一方向的に捉えるモデルから、患者と共有し、関係性を重視する治療的アライアンスへとシフトしていきます。
オタクな話をしよう
本セクションでは、免疫による疼痛コントロールがもたらす可能性のある寄与について、臨床家がより理解するのに役立つかもしれない、現在利用可能な臨床ツールを紹介します。 これらの評価のいずれも、痛みに対する神経免疫ドライバーの特定の状態や、その正確な寄与を直接評価するために検証されたものではありません。 むしろ本セクションでは、特定の臨床所見が、免疫による疼痛コントロールの変化によって生じた機能的な結果をどのように反映し得るのかを探りながら、理論的な神経免疫の概念を臨床実践につなげることを目的としています。
条件付け疼痛調節
Conditioned pain modulation(CPM)では、「痛みが痛みを抑える」という現象を通して、内因性の下行性抑制経路を評価します。 CPMの低下は、下行性抑制コントロールの不足や、神経免疫調節の変化を示している可能性があります。 理論的には、こうした所見は、抗炎症性サイトカイン活性(例:IL-10やTGF-β)の低下、内因性オピオイドを介した鎮痛の障害、調節性T細胞の活動の変化、そして中枢性感作に関与し続けるミクログリアの活性化を反映している可能性があります。
感覚検査
軽い触れ方、チクチクする刺激、温熱刺激などのシンプルな感覚検査で、アロディニアや痛覚過敏が分かることがあります。 これらの所見は、末梢および/または中枢の感作プロセスを示唆している可能性があります。 メカニズムとしては、IL-1β、IL-6、TNF-α、プロスタグランジン、ヒスタミン、NGF、ケモカインなどの炎症性メディエーターが、侵害受容器の作動閾値を下げ、神経の興奮性を高めます。 同時に、ミクログリアの活性化、BDNF放出、ATP-P2X4/P2X7シグナル伝達、そして後角の処理の変化を含む「免疫による痛みの制御」メカニズムが、感覚反応の増幅やアロディニアに寄与し得ます。
圧痛閾値
圧痛閾値(PPT)検査は、痛みを誘発するのに必要な最小の圧力を評価し、侵害受容器の感作や中枢性の痛み増幅に関する間接的な情報を得られる可能性があります。 PPTの局所的な低下は、炎症性サイトカイン、ニューロペプチド、プロスタグランジン、そして機械受容性の侵害受容器に作用するNGFによって媒介される末梢感作を反映していることがあります。 より広範なPPTの低下、特に遠隔の非症候部位での低下は、中枢感作や中枢神経系における侵害受容の増幅(nociceptive gain)の変化を示唆する場合があります。
神経免疫の観点では、びまん性の機械刺激過敏は、持続的なミクログリア活性化、サイトカインによる脊髄後角ニューロンの調節、下行性の抑制経路の機能低下、そして IL-1β、TNF-α、IL-17、ATP シグナル伝達を含む持続的な神経免疫シグナルにより、理論上は反映され得ると考えられます。
時間的加算(テンプラル・サマイション)テスト
時間的加算テストは、同一の刺激を繰り返した際に痛みの知覚が段階的に増えていくことを評価し、脊髄の興奮性と痛みの促通(ファシリテーション)のメカニズムを反映します。 時間的加算の亢進は、後角レベルで起こる「ウィンドアップ」現象の臨床的な関連所見と考えられています。
理論的には、促通性の時間的加算は、持続する侵害受容性入力に加えて、神経免疫による脊髄ニューロンの興奮性増幅が組み合わさることで説明できるかもしれません。 考えられる寄与因子として、ミクログリアの活性化、ATP-P2X4/P2X7シグナル伝達、BDNF放出、サイトカインを介したシナプス伝達の調節、そして抑制性介在ニューロンの機能低下が挙げられます。 IL-1β、TNF-α、IL-6の上昇や、グリア由来のメディエーターは、シナプスの応答性が高まることや、持続的な中枢性感作の維持に寄与する可能性があります。
自己記入式の質問票
CSIのような自己記入式の質問票は、広範な痛みや感覚過敏といった症状を評価することで、ミクログリアの活動を含む中枢の処理の変化に関連し得る神経免疫関連の痛みのメカニズムを、間接的に反映している可能性があります。 ただし、自己申告型のツールであるため、報告バイアスの影響を受ける点には留意が必要です。 それでもなお、痛みが機能や活動、そして日常生活に及ぼす影響を検討するための、役立つ入口になります。
持ち帰りメッセージ
- 痛みは、神経現象だけに限られるわけではありません。侵害受容器、免疫細胞、中枢神経系ネットワークが絶えず相互作用する中で生まれ、動的な「神経免疫システム」を形成します。
- 免疫細胞は痛みの働きを調整する重要な役割を担っており、感作(例:サイトカイン、NGF、ATPシグナル)に関与するだけでなく、抗炎症性および修復促進性のメディエーター(例:IL-10、IL-4、内因性オピオイド、Treg)によって鎮静化(解決)にもつながります。
- 中枢性感作は、脊髄および脳の上位中枢にある回路において、神経免疫の増幅が起こることを指します。特に、ミクログリアの活性化やサイトカイン/BDNF(脳由来神経栄養因子)のシグナル伝達を通じて進行します。
- 神経免疫コミュニケーションは双方向です。免疫メディエーターは侵害受容器に影響を与える一方で、ニューロンは神経ペプチドやケモカインを通じて免疫応答を能動的に調節します。
- QST、PPT、時間的加算、CPM、そして質問票などの臨床ツールは、神経免疫活性そのものを直接測定できない場合がありますが、システムレベルでのその機能的な影響は反映し得ます。
- そのため、理学療法の評価では、痛みの表現型(フェノタイプ)をメカニズムに基づいて解釈することで得られるメリットがあります。つまり、感覚検査を、生物心理社会的な要素や痛みの免疫制御に関するメカニズムに基づいた推論と統合するのです。
- 最終的に、痛みは複雑で適応的、かつ多面的な体験として理解されるべきであり、そのためにはメカニズムに関する洞察と、患者中心の臨床的な統合の両方が必要です。
参考
中枢性感作にとって栄養がいかに重要な要素であるか - ビデオ講義
ヨーロッパNo.1の慢性疼痛研究者ジョー・ナイスによる、栄養と中枢性感作に関する無料ビデオ講義を 見る。 患者がどの食品を避けるべきか、おそらくあなたは驚くだろう!
